As a paleontologist and geologist, time is always on my mind. Nonetheless, such musings do not always connect with millions or billions of years, the so-called “deep time” that earth scientists love to use whenever shocking people who normally ponder shorter time intervals used when, say, measuring the life of a fruit fly, or the length of a cat-themed video.
Still, sometimes other paleontologists and I also try to interpret brief time spans, such as a few minutes, hours, or years, but ones that elapsed millions of years ago. This is where ichnology comes in handy as a tool, as animal traces in particular – such as tracks or burrows – can give “snapshots” of animal behavior in the context of their original ecosystems. For instance, when I look at a limestone layer that was first laid down 95 million years ago and see burrows in that limestone, I think of it as soft, carbonate-laden mud with many small crustaceans digging into it. This is an instance of where imagination becomes a time machine, helping us to create evidence-based explanations that hopefully can be later honed with further scrutiny and re-imagining. When trace fossils are preserved as an assemblage in the sediments of that past ecosystem, whether it was a soil, lake bottom, or beach, the stories can be told in chronological order.
Throw plants into the mix, though, and they can screw up those linear-time stories to the point where you doubt every earth scientist when they tell a story about an ancient land-based ecosystem. Plants can occupy sediments that are hundreds, thousands, or millions of years old, and if their roots penetrate deep enough into these sediments, they may leave both remnants of their tissues and root traces. These geologically fresh root traces then mix with older animal trace fossils, conjuring the illusion of a contemporaneous community, all living happily together. Only a careful examination of the sediment, and which traces cut across which, would help to unravel the real story.
In the preceding video – taken more than four years ago on Sapelo Island on the Georgia coast – I tell such a cautionary tale of what happens when you assume that the animal and plant traces in an old sediment were made at the same time. (Spoiler alert: You would be wrong.)
For more about this relict marsh and the fascinating lessons we can learn from it, please read Fossils In Progress (which includes a short bibliography) and Teaching on an Old Friend, Sapelo Island. Both posts also discuss how to teach students some of these concepts of interpreting fossilization, paleoecology, and geologic time when in the field.
All scientists use tools when investigating how the natural world works. Yet as a traditionally trained field scientist – and an ichnologist – I’ve always been wary of adopting anything more complicated than field notebooks, pencils, tape measures, hand lenses, and cameras. Granted, I did add GPS units to my equipment list starting about 12 years ago and now consider these location-finding devices as standard (and essential) field gear. Still, if you told me even a year ago that I would happily welcome the services of flying robots while tracking alligators on the Georgia barrier islands, I would have smiled and said, “Yes, and Bud Light is my favorite beer.” (Just to clarify: It is not, nor will it ever be.)
Need a better overhead view of barrier-island ecosystems with identified locations, and don’t feel like waiting for the latest satellite photos? I suggest strapping a camera and GPS unit onto a vulture and training it to take pictures while simultaneously recording waypoints. Or, have an aerial drone do the same for you, which will do a much better job, while also not annoying the vulture. (Photograph by Anthony Martin, taken on St. Catherines Island, Georgia.)
So here I am, ready to buy everyone a round of their favorite beverage (perhaps Kool-Aid) in celebration of my being wrong. Earlier this year, an Emory colleague of mine – Michael Page – convinced me that an aerial drone might be a good tool for getting overhead views of ecosystems on the Georgia barrier islands. So as soon as Emory purchased a new, state-of-the-art drone in early 2015, Michael and I plotted to take it to St. Catherines Island for its first real field test in March 2015.
Yeah, I know, it’s not New Horizons, but this drone is still a pretty nifty piece of field equipment, and I’m glad to have added it to my ichnology utility belt. (Photograph by Anthony Martin, taken on St. Catherines Island, Georgia.)
The last time Michael and I were on St. Catherines Island together was two years ago, when we had a group of Emory students help us map gopher tortoise burrows and alligator dens there. (That was fun.) We’ve also been working with a few other colleagues at Georgia Southern University to describe the gopher tortoise burrows and alligator dens on St. Catherines Island over the past few years. So Michael and I figured we could use the drone to aid in this research, starting with the gopher-tortoise burrows.
Perhaps the most persuasive point Michael made about the drone’s potential value was its winning combination of built-in GPS and high-definition video camera. This meant we could instantly map (“georeference”) gopher-tortoise trails between their burrows, as well as the burrows themselves. The latter were easily visible from the wide, white, sandy aprons just outside burrows entrances, and sometimes even show up in satellite photos of the area. The big difference with using a drone versus satellite photos, though, would be in their ‘real-time” capture of these traces – rather than a randomly taken satellite image – while also having much better resolution.
See that hole in the ground? That’s a gopher-tortoise burrow. See those breaks in the grass to the left and right in the foreground, and elsewhere? Those might be trails that connect this burrow to others in the area. How to map all of them? Call in the drone! (Photograph by Anthony Martin, taken on St. Catherines Island, Georgia.)
Can you see gopher tortoise traces from space? Surprisingly, yes. Not only are burrow aprons visible in this GoogleEarth™ photo (denoted by the arrows), but also trails connecting some of the burrows. Although if you find yourself squinting and turning your head sideways to see these, you’ll understand why sending up a drone with a high-resolution camera might be a better way to map these traces. (Image taken from a presentation I gave at the 2011 annual meeting of the Geological Society of America in Minneapolis, Minnesota.)
Most of the gopher-tortoise burrows are in a broad, flat area on St. Catherines that used to be pasture land, but is now being restored to the tortoises’ long-leaf pine-wiregrass ecosystem. This re-located tortoise population has done quite well here, and because of its isolation on St. Catherines, it’s an example of one that does not face as many human-related problems as their compatriots on the Georgia mainland. Its remote location also helped us with trying out the drone, as we didn’t have to worry about it dodging buildings, power lines, or gawking locals, all of which might have complicated its flights.
Almost ready for take-off! Drone pilots/wranglers Alison Hight (left) and Michael Page (right) look for a flat place near a staked gopher-tortoise burrow for setting down our “eyes in the sky.” (Photograph by Anthony Martin, taken on St. Catherines Island, Georgia.)
This was the drone’s maiden voyage on St. Catherines Island, taking off from the gopher-tortoise field. It did just fine. (Video footage by Anthony Martin, taken on St. Catherines Island, Georgia.)
The drone pilots doing a great job, sending the drone around the gopher-tortoise field for a spin. (Photograph by Anthony Martin, taken on St. Catherines Island, Georgia.)
This flight was a big success, in that the drone went up, took lots of video and photos while in the air – all of which was georeferenced – and it came down without crashing. So we decided to try it elsewhere. That’s when we remembered the Atlantic Ocean was only about 500 meters away on the eastern edge of St. Catherines, with a lengthy beach, salt marshes, storm-washover fans, tidal creeks, and a bluff of Pleistocene sand with maritime forest on top of it. So off we went, and we did Flight #2 over the storm-washover fans, salt marshes, and tidal creeks near the north end of the island.
Drones (much like me) operate well in places with wide-open spaces that involve Georgia beaches. Check out how quickly it disappears from view once in the air. (Video footage by Anthony Martin, taken on St. Catherines Island, Georgia.)
Following this flight, we decided to send the drone father north to survey the bluff from just offshore. This was probably the most exciting flight, as we watched it go out to sea, then fly parallel to the shore, with its camera trained on the coastline.
Michael setting down the drone on a almost-flat surface as Alison prepares it for take-off. The yellow yardstick serves as an easily visible scale that can be used to estimate ground-level distances. (Photograph by Anthony Martin, taken on St. Catherines Island, Georgia.)
Off we go, into the wild blue yonder. (Video footage by Anthony Martin, taken on St. Catherines Island, Georgia.)
Bringing it back home. Look for the spot near the top-center of the photo for our “hand lens in the sky.” (Photograph by Anthony Martin, taken on St. Catherines Island, Georgia.)
Coming in for a soft landing, which is much preferred over the other type of landing. (Video footage by Anthony Martin, taken on St. Catherines Island, Georgia.)
So following these inland and coastal successes, which clearly were applicable to studying gopher tortoises and coastal geology, it was time to try using the drone to look at the apex predators of the island – alligators – and their traces. The next day,while scouting areas further to the south for alligator dens and tracks, we paused on a causeway cutting through a salt marsh. Because the marsh was at low tide, its mudflats were exposed, which allowed a few big animals to walk across it and leave their tracks, and for us to see these tracks.
At least two of the trackways were from alligators, made distinctive by their sinuous tail drags, arcing footprints, and belly drags. I suspect the other trackways were from feral hogs, but I couldn’t tell for sure because they were in squishy mud beyond my carrying capacity. Which is to say, I would have quickly immersed myself in this environment had I gone any further out. Gee, if only we had some way to photograph those trackways from above, better helping us to see their lengths, patterns, and directions.
A salt-marsh mudflat at low tide, with low marsh and a patch of forest (hammock) in the background. See the alligator trackway to the left, where the alligator turned? Look in the middle and you’ll see two more trackways that are probably from feral hogs, and another curving trackway to the right that is from another alligator. (Photograph by Anthony Martin, taken on St. Catherines Island, Georgia.)
Why wade into waist-deep salt-marsh mud to track an alligator when you can stay safely (and cleanly) on dry land, telling a drone what to do? (Photograph by Anthony Martin, taken on St. Catherines Island, Georgia.)
So it was time for another flight, and the drone’s first alligator-track-mapping mission, which I’m pleased to say was a success. One example of that success is conveyed by the following photo, which made me gasp when I first saw it. There were the two alligator trackways and the two hog trackways, but also two not-so-clear trackways I had missed and a clear view of where the hogs had dug along the marsh edge. This photo similarly evoked a collective “Ooooo!” when I showed it to an audience the next week at the Southeastern Section meeting of the Geological Society of America meeting in Chattanooga, Tennessee. My talk was a progress report on the alligator dens of St. Catherines Island, but I threw in this photo toward the end of it to show how drones might help with some of our tracking alligator movements through difficult-to-access environments on the island.
OK, you’re probably wondering by now how good those photos and videos taken by the drone might be, and whether or not any useful science can come from them. See that guy in the lower center of the photo? That’s me, pointing to each of the two alligator trackways, with the yellow yardstick providing an additional scale to the left. Notice also the probable feral hog trackways in the middle and fainter ones to the right, as well as the “hogturbation” (rooting disturbance caused by hogs) in the upper left of the photo. As an ichnologist, I was pretty darned pleased by this picture, and I want more like it. (Photograph by The Aerial Drone, taken on St. Catherines Island, Georgia.)
Lastly, I was also happy to see that drones have their own ichnology, in that they make flight traces. I’ve been long fascinated by flight traces – called volichnia by ichnologists – and have done my best to describe these in modern birds of the Georgia coast, as well as bird flight traces in the fossil record. Given the right substrate, anatomy, and behavior, the take-off and landing traces of birds and other flighted animals can preserve well enough for us to interpret them for their true nature.
Now, to do the same for a drone requires knowing how they have vertical take-offs and landings, using rapidly moving rotors. This means air will be pushed down onto the substrate directly underneath the drone, then dissipated abruptly outside that zone. The result would be a sem-circular depression slightly more that the maximum width of the drone, and one that would look very much the same whether made by a take-off or landing. The difference would be in the timing of the landing-pad traces: if obscured by the depression, then it was taking off, but if they are impressed on the depression, then it was landing.
Drone coming in for a landing, already pushing aside pine needles on the forest floor and making its landing trace. (Photograph by Anthony Martin, taken on St. Catherines Island, Georgia.)
Drone landing trace, minus the drone. Do you see the square pattern in the middle of the oval depression? That’s the outline of the drone, defined by its landing gear.(Photograph by Anthony Martin, taken on St. Catherines Island, Georgia.)
So now we know that a drone can be used for conservation biology, coastal geology, behavioral ecology, and – most importantly – ichnology. How about art? Yes indeed. Once we got back to the Emory campus, Michael handed over the footage to Steve Bransford, a skilled videographer employed by Emory and founder of Terminus Films. Given all of the drone footage, he snipped out the boring parts (always a good thing to do), added a few maps at the start to orient the viewers, put in a soothing soundtrack, and basically created an aesthetically pleasing and extraordinarily educational video. So we submitted it for consideration as an video in the peer-reviewed online journal Southern Spaces, which was founded at Emory University. Much like an aerial drone on an unobstructed coastline, it sailed through peer review and is now available for viewing by all who have an Internet connection.
St. Catherines Island Flyover from Southern Spaces on Vimeo. Never mind the stern message: just click on the link or the video and it will play. Once it does start playing, please watch it on a big screen, sit back, and enjoy the ride. Also be sure to read the accompanying article linked to the peer-reviewed online journal Southern Spaces.
What’s aerial adventures await us next? We’ll see, as we have plenty of visual information and data to process from our previous visit. But for now we can be pleased to have shown the value of an aerial drone as both a scientific instrument and a means for engaging our senses with soaring imaginations.
Acknowledgements: Many thanks to the St. Catherines Island Foundation for its support of our research on St. Catherines, and to Royce Hayes and Michael Halstead for their assistance on field logistics. We also appreciate the expert piloting of the drone by Alison Hight while on St. Catherines. Steve Bransford did a fantastic job with creating the video for the Southern Spaces article, which should win the Georgia equivalent of an Oscar. Input from the editor of Southern Spaces, Allen Tullos, improved our article accompanying the video, and we are grateful to the staff of Southern Spaces for their quality service in putting this video and article online. And as always, many thanks to Ruth Schowalter for her help and support, in and out of the field.
(Author’s note; The following post is a republished article of mine, originally published on June 12, 2014 by The Conversation and later republished by The New Republic, The Guardian, Quartz, and several other online news sources. However, this post is an embellished version, in which I include a paragraph on dinosaur microbiomes omitted from the original, and it uses my personal photographs and captions to illustrate its points about dinosaur paleoecology. So you might say this is the “director’s cut.” Many thanks to The Conversation editor Nick Lehr for helping turning my rough prose for the original article into one more readable for a general audience.)
Like many moviegoers this summer, I plan to watch Jurassic World. And because I’m a paleontologist, I’ll cheer for the movie’s protagonists (the dinosaurs) and jeer at the villains (the humans). But no matter how thrilling this movie may be, one question will plague me throughout: where are the dung beetles?
This mural depicts theropod dinosaurs (foreground) and sauropod dinosaurs (background) as part of a Late Jurassic ecosystem about 150 million years ago. OK, so this ecosystem has some producers (plants), primary consumers (herbivores, the sauropods), and secondary consumers (carnivores, the theropods). What’s missing from this picture that would be needed to make this a real, functioning ecosystem? If you said “Dung!” and “Dung beetles!,” you’re on the right track. (Mural by Robert F. Walters and Tess Kissinger (Walters & Kissinger) at the Carnegie Museum of Natural History, photograph by Anthony Martin.)
Dung beetles – which are beetles that eat and breed in dung – would be only one of many ecological necessities for an actual Jurassic World-style theme park. Yes, cloning long-extinct dinosaurs is impossible. But even if dinosaur genomes were available, the animals couldn’t simply be plopped anywhere.
So for the sake of argument, let’s say an extremely wealthy corporation did manage to create a diverse bunch of dinosaurs in a laboratory. The next step in building a Mesozoic version of Busch Gardens would be figuring out how to recreate – and maintain – the dinosaurs’ ecosystems. Accomplishing this goal would require a huge team of scientists, consisting (at minimum) of paleontologists, geologists, ecologists, botanists, zoologists, soil scientists, biochemists and microbiologists.
Such a team then would have to take into account countless interacting factors for the dinosaurs’ recreated habitats. And perhaps they could take a page from rewilding efforts that are currently taking place throughout the world.
In a memorable scene from the original Jurassic Park, paleobotanist Dr. Ellie Sattler examines an impressive heap of an ill Triceratops’s feces to look for digested remains of a toxic plant.
One of my favorite scenes in Jurassic Park (1993), when Dr. Ellie Sattler (played by Laura Dern) affirms her Ph.D. (= “Piled Higher and Deeper”) by unhesitatingly plunging her hands into a massive pile of Triceratops feces. Please note her sensible footwear, suitable for running away from theropods planning to add her to the local food web.
Here, the filmmakers touched on a key challenge for recreating an environment from a different geologic period. Many modern plants have evolved defenses against herbivores, which include toxins that can swiftly impair any animal that hasn’t adapted to them. Consequently, a time-traveling Triceratops would be taking a big risk with every visit to its local salad bar.
Paleobotanists could try to solve this problem by cataloging fossil plants that lived at the same time as plant-eating dinosaurs, before picking out descendants of those plants that are still around today. Still, plant lists will never be good enough to say whether or not a Triceratops, Stegosaurus, or Brachiosaurus ate those plants or if they could eat their descendants.
The same might hold true for carnivorous dinosaurs, which – for all we know – may have been picky eaters. For instance, although some Triceratops bones hold tooth traces of Tyrannosaurus, there’s no way to be sure a genetically engineered Tyrannosaurus would eat an equally inauthentic Triceratops (even if it were organic and free-range).
Did tyrannosaurs ever eat Triceratops? Oh yeah, and with gusto. Tooth trace fossils in Triceratops hip bones (red arrows) happen to match the dental records of Tyrannosaurus rex, which lived as the same time (Late Cretaceous, 65-70 million years ago) and place (western North America) as Triceratops. Also think about how much meat was covering that hip bone, which means the Triceratops must have been dead when this tyrannosaur was helping to recycle its body into the ecosystem. (Specimen in Museum of the Rockies and photograph by Anthony Martin.)
Yet another food-related dilemma is that we also are not quite sure how most dinosaurs digested what they ate. For instance, many modern animals – from termites to humans – require a suite of gut bacteria to break down and assimilate nutrients from food. Even if microbiologists somehow successfully recreated the microbiome of a dinosaur, how would you prevent it from acquiring modern gut parasites? Dinosaur coprolites (fossil feces) tell us that some dinosaurs had gut bacteria and parasites: but how to engineer the right bacteria and exclude the wrong parasites?
So despite a century of dinosaur flicks portraying tyrannosaurs and other predatory dinosaurs gratuitously munching humans, one bite of our species – or other sizable mammals – might make them sick. In other words, there’s no accounting for taste.
The lack of dung beetles in that same scene with Dr. Sattler also may have explained why the Triceratops’s feces were piled so high. We know from fossil burrows in dinosaur coprolites that dung beetles fed on dinosaur droppings at least 75 million years ago. Similarly, Late Jurassic dinosaur bones from nearly 150 million years ago hold the traces of carcass-eating insects.
A large, 75-million-year-old coprolite – attributed to the hadrosaur Maiasaura – filled with digested plant debris, but also with dung-beetle burrows. One burrow is sliced length-wise and runs diagonally (upper right to lower left), and another is in cross section and pointed toward you (upper right). Specimen is from the Museum of the Rockies but was part of a traveling display at Fernbank Museum of Natural History in the late 1990s. (Photograph by Anthony Martin.)
Late Jurassic (about 150 million-year-old) dinosaur bone with insect borings, which are credited to carcass- and bone-eating insects that used these bones for food or breeding soon after the dinosaur was dead. Specimen on display at Dinosaur National Monument near Vernal, Utah. (Photograph by Anthony Martin.)
This makes sense: wastes, bodies and other forms of stored matter and energy must be recycled in functioning modern ecosystems. Accordingly, to maintain the productivity of these dinosaurs’ ecosystems, animals that perform essential services to the ecosystem would need to be introduced. These include pollinators, such as bees, beetles and butterflies, as well as seed dispersers, like birds and small tree- and ground-dwelling mammals. Thus Masrani Global – the imaginary corporation tasked with creating Jurassic World – should have added entomologists (insect scientists), ornithologists and mammalogists to the career opportunities page on its mock website.
Can we learn anything useful from such fanciful reconstructing of long-gone ecosystems, where large animals once roamed? Sure. In so-called “rewilding” projects, imagination meets real science. These projects, which attempt to restore ecosystems by closely mimicking their previous iterations, often include reintroducing locally extinct animals.
Perhaps the most famous and successful of such rewilding projects began just after the release of the original Jurassic Park. In 1995, wolves were reintroduced to Yellowstone National Park. Although admittedly not as exciting as releasing a pack of velociraptors into the woods, the reintroduction of wolves – which had been extirpated from the area earlier in the 20th century – had a dramatic restorative effect.
If you looked for these tracks in Yellowstone National Park before the original Jurassic Park came out in 1993, you would have been disappointed. They’re from gray wolves (Canis lupus) and are signs of a now-thriving population of these apex predators reintroduced to the Greater Yellowstone Ecosystem in 1995, which has since caused big changes there. (Photograph by Anthony Martin.)
After the wolves gorged on elk – which, without predators, had overpopulated the region – riverine foliage grew more lushly. This prevented erosion and expanded floodplains, which gave beavers a better habitat to get to work damming rivers. A similar experiment is taking place in Europe, where increased numbers of large carnivores, such as wolves, bears and lynxes, are reshaping their ecosystems closer to their original states.
Bolstered by these successes, rewilding proponents have even proposedreintroducing elephants, lions, cheetahs and other animals to parts of North America as ecological proxies to mammoths, American lions and American “cheetahs” that lived only a little more than 10,000 years ago in those areas. Given the much shorter elapsed time since their extinction, enough similar species today and no need for genetic engineering, a “Pleistocene Park” – Pleistocene being the geological epoch that was about 2.5 million to 11,700 years ago – would be far easier to achieve than a Jurassic World (while also being more alliterative).
You want a “Pleistocene Park”? Here’s a start, with herds of large primary consumers (Bison bison, otherwise known as “bison”) and grasslands in Yellowstone National Park, which overlap in range with secondary consumers wolves and grizzly bears. Now just add some elephants, lions, cheetahs, and a bunch more dung beetles, and you should be set. Wait a minute: you say the National Park Service wouldn’t approve of that? Oh well, one step at a time. (Photograph by Anthony Martin.)
So to any corporations out there that are thinking of making such a park, do us a big favor: whatever you do, don’t forget to include dung beetles.
Let’s try a science-education experiment. Give a child a live clam and ask, “Can this animal fly?” and I predict her or his answer – accompanied by much giggling – will be “No!’ But if you ask, “Can you fly?”, the answer may change, especially if this child has already flown on an aircraft. So of course humans can fly, but to do this, they require machines, paragliders, or other technological aids in order to move through the air and – this is important – arrive on the ground safely.
For clams that try to fly, they end up with more than shattered dreams. How did these clams (Mercenaria mercenaria, also known as quahogs or “hard clams”) end up doing Humpty-Dumpty impressions on a wooden pier? Please read on. (Photograph by Anthony Martin, taken on Jekyll Island, Georgia.)
In a similar way, clams can fly. They just need a little help from other animals that can fly and willingly give them a temporary lift from the earth they and their molluscan relatives have known for all of their evolutionary history. Compared to most of our forays into the air, though, these flights are much more limited. Clam aerial exploits are brief and mostly vertical, with little time for them to appreciate the view from above or otherwise experience unusual sensations. They go up, then they come down, and fast.
Clams do not have landing gear. So they can hit the ground hard, especially if their free fall happened after a lengthy trip up into the air and the ground surface is hard: think of a sandflat at low tide, a paved parking lot, or a wooden boardwalk. A a result, the most common end to clam flights is a shattered shell, which is quickly followed by the demise of the clam as it is consumed by the very same animal that bestowed it with flight, however brief and self-serving.
Traces of a unidirectional vertically oriented clam flight (otherwise known as “falling”) that did not end well for the clam, but worked perfectly for the flying animal that took it for a ride. Notice the impact trace on the hard sandflat, outlining the ribbed shell of the clam (probably Dinocardium robustum) and bits of shell. Most of the probably-still-alive-but-definitely-dying animal was dragged off to a nearby spot so that its soft parts could be eaten by the same perpetrator that took it for a ride. (Photograph by Anthony Martin, taken on Sapelo Island, Georgia.)
So just what flying animals do such dastardly deeds, taking hapless clams up for a ride, only to drop them to a certain death? By now the gentle reader has probably figured out birds are responsible for this blatant bivalvicide, and some may have already known that seagulls are the most likely culprits. In some coastal areas and during low tides, some seagulls fly over exposed sandflats and mudflats, searching for the outlines of clams buried below the surface. These avian ichnologists then swoop down, land, pick up the clam with their beaks, take off, and then once high enough, they drop them, serving up instant raw clam on the half (or quarter, or eighth) shell. Typically all that is left is a jigsaw puzzle of clamshell pieces and the seagull perpetrator’s footprints, but with the latter only evident on muddy or sandy surfaces amenable to preserving tracks.
Ichnological evidence of who killed the clam, provided by the tracks a laughing gull (Larus altricilla).The other half of the shell was broken by its falling onto the sandflat elsewhere, then the gull carried its clam on the half-shell to a more scenic place for its meal. (Photo by Anthony Martin, taken on Little St. Simons Island, Georgia.)
I found this behavior so compelling that I started my book Life Traces of the Georgia Coast (2013) with a story about a laughing gull (Larus altricilla) and the traces of its unwitnessed predation on an Atlantic cockle (Dinocardium robustum), seagull behavior on the Georgia coast. I was not the first person to note this method of clam-smashing by seagulls, as it has been documented by other scientists in parts of the U.S. and abroad, and has been caught on video. Amazingly, though, despite more than 15 years of visiting the Georgia coast, I had never actually witnessed seagulls dropping clams. instead I had only performed post-mortem forensics, in which I would find broken clamshells on hard sandflats accompanied by seagull tracks, telling tales of murder most fowl.
Video footage of a western gull (Larus occidentalis) picking up a clam, flying up about 10 meters (> 30 feet), and dropping it onto rocks to crack it open. After this doesn’t work the first time – and after shooing away a potential clam-stealing rival – it tries again, and is presumably successful. It’s almost as if this gull is using a scientific methodology, isn’t it? (The videographer is only credited as ‘Trisera’ on the YouTube page, and I don’t know where it was filmed, but suppose it’s on the western coast of the U.S.)
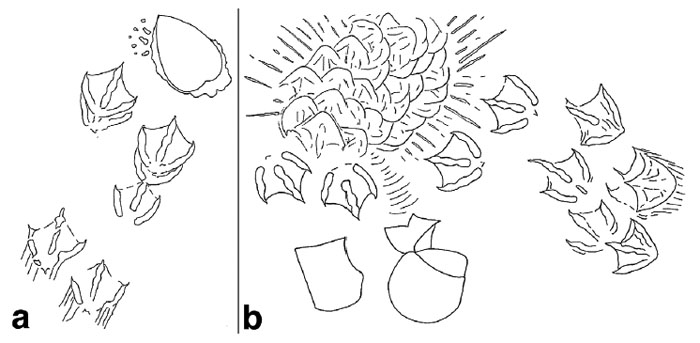
Here’s the first illustration a reader will see in my book, Life Traces of the Georgia Coast (2013, Indiana University Press), which I drew to provide a visual forensic analysis of how an Atlantic cockle met its demise at the hands of – er, I mean, wings and bill of – a laughing gull. Part (a) depicts the gull landing after recognizing the outline of the cockle from the air, stopping, and extracting it from the sandflat. Part (b) shows where the cockle was dropped and broken successfully, accompanied by the gull landing and trampling the area as it enjoyed its clam dinner.
This meant I was more than overdue to get visual confirmation of gulls killing clams, which was finally granted just a few weeks ago during a recent trip to Jekyll Island (Georgia). It was the day after I had given an invited talk at the annual meeting of The Initiative to Protect Jekyll Island environmental group, and while my wife Ruth and I were relaxing before leaving the island, but of course were also observing whatever nature we could.
In that spirit, and while sitting on a deck on the west side of the island and looking at a mudflat (in between swatting sand gnats), we noticed a seagull flying about 10 meters (>30 feet) above a wooden pier. At one point, it paused its ascent, and we saw an object fall from its mouth and down toward the pier. Thunk! We clearly heard the impact of the object correlate with what we saw, and with much excitement realized that we had just witnessed seagull clam-cracking for the first time.
A mudflat replete with mud snails (probably Ilyanassa obseleta), grazing away and making gorgeous meandering trails on the western side of Jekyll Island (Georgia). But wait, what are those big white chunks on the same surface?
Why, look at that: hard clams (Mercenaria mercenaria) in an unnatural state, i.e., disarticulated, broken, and dead on the surface of the mudflat. These clams normally burrow into and live under the mud, and usually manage to stay intact if they stay below the surface. The pieces of clams here must have bounced off the wooden pier, which is casting a shadow in the lower right-hand side of the picture. (Both preceding photographs by Anthony Martin and taken on Jekyll Island, Georgia.)
What was most surprising to me about this broken-shell assemblage on the pier was how it was represented only by the hard clam, or quahog (Mercenaria mercenaria). These thick-shelled clams are quite common in sparsely vegetated muddy areas of salt marshes, burrowing into the mud and connecting their siphons to the surface so that they can filter out suspended goodies in the water during high tides. During low tides, however, they become vulnerable to avian predation. Despite being “hidden” in the mud, somehow the seagulls spotted them from the air, landed next to them on the mudflat, and pulled them out of the mud. They then used the nearby pier as an anvil, and the clam’s hard, thick shell unwittingly became its own hammer when they hit the pier after falling from a fatal height.
The horror, the horror: a clam killing “ground,” thoughtfully supplied by humans for seagulls in the form of a long, hard, wooden pier. (Photograph by Ruth Schowalter and Yours Truly for scale, taken on Jekyll Island, Georgia.)
OK, now it’s time to think about broken clams and deep time. If you found such an assemblage of broken shells of the same species of thick-shelled clams in a geologic deposit, how would you interpret it? Would you think of these broken shells as predation traces, let alone ones made by birds? Which also prompts the question, when did seagulls or other shorebirds start using flight and hard surfaces to open clams? Did it evolve before humans, and if so, was it passed on as a learned behavior over generations as a sort of “seagull culture”?
All of these are good questions paleontologists should ask whenever they look at a concentration of broken fossil bivalves that are all of the same species, and overlapping with the known geologic range of shorebirds. In short, these may not be “just shells,” but evidence of birds using gravity-assisted killing as part of their predation portfolio.
I’ve always been a big fan of aquariums. Having grown up in the landlocked Midwest and not seeing an ocean with its bountiful life until I was 20 years old, I am still drawn to the old-school charm of big tanks filled with salt water and populated by exotic fish and other sea critters. These environments, however artificial, never fail to inspire awe and wonder. Even better, they often teach me something new and relevant each time I pay closer attention to what they hold.
A seahorse, of course, is not a horse. But that’s not the only way seahorses differ from horses, in that they leave trails instead of tracks. Intrigued? Yeah, me too. (Photograph by Anthony Martin, taken at the UGA Aquarium, Skidaway Island, Georgia.)
Nonetheless, I also have a “problem,” which manifests itself whenever I’m at an aquarium, walking along a beach, sitting on a park bench, driving down a road, or, well, conscious. As an ichnologist, I’m constantly looking for animal traces. Then once found, I study these traces carefully so that they may inform me whenever I see similar traces in the fossil record. But because I’m a land-dweller and rarely have the opportunity to snorkel or scuba-dive, aquariums come in handy for observing traces of aquatic animals I might not often see. Particularly helpful are aquariums in which the people caring for them were kind enough to include sand on their bottoms (the aquariums, that is).
So last weekend, while leading a class field trip to the Georgia coast and after a wonderful boat ride to Wassaw Island and back, I eagerly joined my students in viewing a salt-water aquarium. This particular venue was the UGA Aquarium (UGA = University of Georgia, Athens) is maintained by the UGA Marine Extension Service (MAREX) on Skidaway Island, Georgia. Our visit was especially satisfying because we were there on a Sunday afternoon, when the aquarium is closed to the public. This luxury afforded us plenty of room and quietude, qualities that are rumored to enhance learning.
Within just a few minutes of entering the main room, one tank to the right caught my eye, and not just because of its pretty colors, but for its denizens and traces on the sandy bottom of that tank. It contained seahorses, fishes that are so odd compared to other fishes, we humans had to compare them to hoofed domesticated mammals. The best part of all, though, was that this tank had lots of intersecting grooves and circular imprints on its sandy surface, which no doubt had been made by the seahorses.
A seahorse (Hippocampus sp.) showing off its lack of swimming skills by moving along the sandy bottom of a tank. Gee, what are all of those meandering and intersecting grooves in the sand and circular imprints? I wonder what made those? Sorry, first guess doesn’t count. (Photograph by Anthony Martin, taken at the UGA Aquarium, Skidaway Island, Georgia.)
All seahorses are under the genus Hippocampus, which consists of more than fifty species. Evolutionarily speaking, they are ray-finned fish (actinopterygians) and share a common ancestor with pipefish and sea dragons (Sygnathidae). The oldest known fossil seahorses are in Miocene Epoch rocks, from about 13 million years ago. Besides their equine-like profiles, they are well known for their prehensile tails, which can either grasp onto algae, sponges, or corals, or curl up underneath them as they swim.
However, seahorses are never going to inspire bets at underwater race tracks, as they are among the slowest-swimming of fish, propelled mostly by tiny pectoral fins while moving upright. Still, they don’t need to be fast, as they are very successful predators, with about 90% accuracy in nabbing fast-swimming small crustaceans that get too close to their mouths. Seahorses also don’t need to swim away from larger predatory fishes that might wish to pick them from a seafood menu. Whenever seahorses attach to algae and corals, they sway in harmony with their temporary hosts, effectively blending in with their surroundings.
One point I keep in mind whenever visiting an aquarium, zoo, or other such enclosures is how these can alter so-called “normal” behaviors of their animals. In this instance, the smaller space of this tank, combined with little material for attachment, meant these seahorses were more likely to swim along its bottom then they might in an open ocean. Accordingly, they had made lots of traces in the sand: mostly undulating grooves, but a few circular impressions from their curled tails plopping onto one side or the other.
A seahorse making tail trails while swimming along the bottom of an aquarium. Notice how the trail would become less linear, wider, and more circular if the tail flops over to one side or another, involving a greater area of the curled end. (Photograph by Anthony Martin, taken at the UGA Aquarium, Skidaway Island, Georgia.)
A close-up of those trails left by swimming seahorses dragging their tails along a sandy surface. Also, check out the overlapping circular “plop” traces on the right, made by the curled part of the tail? (Photograph by Anthony Martin, taken at the UGA Aquarium, Skidaway Island, Georgia.)
What’s the take-home message of these observations for ichnologists, geologists, and paleontologists? That experience matters, as does questioning preconceived notions about what we might observe from the geologic record. Take a look at the preceding photo, and tell me – quite honestly – that your very first interpretation of the tracemakers would have been “fish,” let alone “seahorse.” Instead, I think nearly everyone (yes, me too) would have reached for the easiest answer, which would have been “worm trails,” similar to how geologists reflexively apply “worm burrows to anything small, tubular trace fossil they encounter at an outcrop. Wrong, wrong, wrong.
So next time when looking at rocks formed in marine environments – whether from the last 13 million years or much older – and these rocks host lots of “worm trails” on their surfaces, ask yourself who else could have made such trails, and how. Reach beyond easy and ordinary explanations, and imagine. Oh, and when you go to aquariums, don’t just look at their sea-life, but also the traces of the sea-life in them.
Despite more than 15 years of visiting the Georgia coast, studying its traces, and taking students on field trips to its barrier islands, I always marvel at how each trip is different, bestowing new insights and lessons to both me and my students. So a trip there this past weekend was no exception, and perhaps the most intriguing phenomenon I encountered during it was of some “mere” scrapings in a sandy road on Wassaw Island, Georgia.
Who needs a Mystery Date when you can have a Mystery Trace? Here we have some enigmatic scrapings in a sandy road on Wassaw Island, Georgia. What could have made these, and why? (Photograph by Anthony Martin; scale in centimeters.)
Wassaw Island is a National Wildlife Refuge, and I’ve mentioned it before as the one island of the Georgia coast that most closely approaches the ideal of “pristine,” a label blithely applied to nearly any Georgia barrier island regardless of how much humans had modified their landscapes. Current estimates are that it Wassaw only about 600 years old, which means that Native Americans had barely populated it by the time the Spanish arrived in the 16th century. Thus whenever I teach my biannual Barrier Islands class, I like to include a field trip to Wassaw Island so my students can appreciate the close-to-natural state of its ecosystems. We then contrast their experiences there by visiting overdeveloped Tybee Island on the same weekend, giving my students the opportunity to think about “before and after” conditions of Georgia barrier-island ecosystems.
Even better for my students, our leader for the field trip to Wassaw was not me, but John “(“Crawfish”) Crawford, one of the most knowledgeable naturalists on the Georgia coast. Employed by the University of Georgia Marine Extension Service on Skidaway Island, John regularly takes groups on an open boat to Wassaw Island for day trips. These trips never disappoint for the sheer variety and richness of natural history learned along the way, whether on the boat trip there and back, or on the island itself. I’ve been to Wassaw four times with John as a guide, and each time with him have seen something novel there. (I mean, how often do you see a decapitated seagull?)
Into the Woods, Wassaw Island style! With our intrepid guide (John “Crawfish” Crawford) leading the way into the maritime forest of Wassaw, my students were in for a world of discovery on this beautiful Georgia barrier island. (Photograph by Anthony Martin.)
Just one example from this most recent trip I’d like to share are traces I have never before seen, or, more likely, never before noticed. We encountered it while walking down a sandy road on Wassaw used more often by deer and alligators than humans. The traces were systematic and widespread scrapings of the top few centimeters of the road, some of which resolved themselves as curved to linear features with finer grooves in their interiors. Because they did not match the feeding traces of feral hogs (Sus scrofa) or nine-banded armadillos (Dasypus novemcinctus), I was intrigued. Who made these, and why?
Just in case you missed it the first time, here’s that photo again. Yes, this will be on the exam: why do you even ask?
A closer look revealed the traces were overlapping sets pf five evenly spaced grooves, corresponding with five thin-fingered hands. These could only belong to the most dextrous, industrious, and resourceful denizens of maritime forests and other environments on the Georgia coast, raccoons (Procyon lotor). When I queried John about these traces, he confirmed that not only were they made by raccoons, but also were a result of their “mining” the sand. The raccoons, using their front paws, methodically raked the loose sand to expose shallowly buried acorns dropped by the many old and mighty oaks lining the road, indulging in an all-you-can-eat acorn feast.
Close-up of the same mystery trace seen in the previous photo, but this time more groovy. Check out the curving, parallel set of five grooves (left) and the partial track (right), telling us that a masked bandit left its mark. (Photograph by Anthony Martin; scale in centimeters.)
Although raccoons are infamously omnivorous, in winter months they depend on acorns for much of their diet. Thus considering that the Georgia coast was still in winter, and that a sub-freezing cold front had just passed through the area a few days before, it was not surprising to see this evidence of extensive acorn foraging.
OK, time to replace my floppy coastal-geologist hat with my more stylish paleontologist hat to ask this question: Would such traces preserve in the geologic record, and if so, would they be recognizable for both the tracemaker (raccoon) and behavior (foraging)? Probably not for both, as the loose quartz-rich sand in a maritime forest would have few chances of being buried intact and cemented in a way that would “freeze” the details needed to discern both tracemaker and its intent. Yet these traces would lend some insights to interpreting disturbed zones in the upper parts of fossil soils, especially those that might have preserved acorns or other nuts in them.
So next time you’re in a maritime forest during the winter and come across some odd scrapings in the road, take a closer look and ask yourself a few questions about them. Who made them? Why did they make them? How do these traces relate to the broader ecology of the area? Would they be preserved in the fossil record, and if so, could we properly interpret them? Then ask yourself what you’ll find next time you go to the same place and look just a little bit closer.
The wide variety of modern bird behaviors – as well as the traces that result from these behaviors – continue to captivate and fascinate me. Given recent revelations of birds’ dinosaurian ancestry and the interrelationships of modern birds (an evolutionary history spanning more than 150 million years), this wonderment should be expected. Accordingly, then, the traces made by modern birds can be equally varied, and can serve as guides to the behaviors of their predecessors, especially when made by birds interacting with ecological margins (ecotones).
A mixture of tracks left by boat-tailed grackles (Quiscalus major) and American crows (Corvus brachyrhynchos) on wind ripples in the upper part of a sandy beach. So if paleontologists found something similar in the geologic record, would they be able to say more than “Looks like a bunch of birds were walking around”? That’s why we look at modern traces and their associated behaviors: to get beyond such easy (and terribly incomplete) answers. (Photograph by Anthony Martin, taken on Tybee Island; pen is about 15 cm (6 in) long.)
The most recent example I witnessed of bird tracemakers and their traces in an ecotone setting was last month on Tybee Island (Georgia). Tybee is a barrier island just east of Savannah, and one I had visited in May, when I noted burrowing wasps in the coastal dunes there. The tracemakers were boat-tailed grackles (Quiscalus major), a passerine bird (“songbird”) that people commonly see and hear along the Georgia coast. Grackles belong to to an evolutionarily related group (clade) called Icteridae, colloquially known as “blackbirds.” I frequently see grackle tracks on the upper parts of beaches and in the dunes, where they are oftentimes the most common vertebrate traces above the high tide mark on Georgia shorelines.
What was strikingly atypical this time, though, was how all of the grackles I saw making tracks were adult females. Female grackles are distinguished from males by their brown coloration, whereas the males are iridescent black, almost deep purple when viewed in the right light. Female adult grackles are also noticeably smaller than adult males, at about 70% their lengths and half their weights. Like most passerine birds, grackles have four-toed anisodactyl feet, with the “thumb” (digit I) pointing directly backwards with respect to its three forward-pointing toes (digits II-IV). Such tracks show their feet are well adapted for grasping branches in trees; yet they hang out along shorelines and nest near water bodies. I also wondered whether the tracks of this gender-sorted assemblage could be distinguished from those of the larger males, but didn’t get a chance to test this idea.
Girlfriends going out for a bite to eat by the beach: a group of boat-tailed grackles – all female adults – foraging in between the sea oats on the south end of Tybee Island. Here they were on the seaward side of the dunes, and just before sundown. (Photograph by Anthony Martin.)
Yet it wasn’t track sizes that caught my attention: it was what they were doing and the traces they were leaving. They were actively foraging, walking in between stalks of the sparsely populated sea oats (Uniola paniculata), which were barely holding down the dunes. This meant lots of slow, methodical walking with their heads down, and beaks actively snatching anything of interest. What were they finding and eating? On an over-developed island like Tybee, it could be almost anything. Grackles are notoriously omnivorous, which explains why they’ve easily adapted and thrived along the eastern coast of the U.S. despite extensive human alterations to this island and elsewhere.
Grackles in different feeding postures: two with their heads down and feet together (foreground and right), and another with her head up and left leg ahead of her right, and all after walking slowly and stopping often. With that in mind, think of the trackway patterns that would correspond with these movements and postures. (Photograph by Anthony Martin, taken on Tybee Island.)
So here’s what’s cool: these grackles were eating locally by chowing down on sea oats. That’s right, given all of the human-provided junk food they had available, they were going for the all-natural, organic, raw, and totally vegetarian option. (Tragically, it was not gluten free. But I think they were OK with that.) As a result, their tracks showed lots of short steps (diagonal walking) punctuated by “T-stops,” where they stopped to place their feet side-by side (making a “T” pattern), all of which were accented by beak traces, the last of these intersecting depressions formerly occupied by the sea-oat grains.
A close-up of the grackle from the previous photo, showing exactly why it stopped it with its feet together and put its beak down to the sand: fallen grains of sea oats. (Photograph by Anthony Martin, taken on Tybee Island.)
Another close-up of a grackle, but with sea oats in her beak. More importantly, check out the tracks behind her, the little depression where the oats laid on the sand (arrow), and the beak mark next to it that she made just before grabbing the grains. (Photograph by Anthony Martin, taken on Tybee Island.)
Boat-tailed grackle tracks that say, “I’m out looking for food, and whole grains only, please.” Note the “T-stop” pattern in the tracks and a beak impression within the trackway (center bottom) coinciding with some sea-oat grains, and a similar set of traces toward further down the trackway. (Photograph by Anthony Martin, taken on Tybee Island.)
So if you’ve read anything written by me before now, you probably know what I’m going to do next. (No, not that. But maybe next time.) I’m probably going to say, “Hey y’all, why don’t you look for traces like these next time you’re out walking along the beach?” But I’m also likely to say, “Gee, I wonder if traces like these would show up in the fossil record?” Both are important questions to keep in mind, even though the first deals with the here and now, whereas the other dives into deep time.
As a paleontologist, though, I’m all about the deep-time question. For example, when did the ancestors of grackles and other blackbirds start eating seeds from the ancestors of the sea oats, and in coastal environments? How would we know when these proto-grackles started having cereal for breakfast? If any trace fossils that look like the ones shown here do somehow got preserved, they should help connect those dots between all of the genes, bones, and other scientific evidence we use to figure out the evolution of this diverse clade of blackbirds.
Yeah, I know, it’s a body fossil. But hey, it’s the Berlin specimen of Archaeopteryx, probably the most famous body fossil in the world, so it’s OK. I was lucky enough to see it in person at the Museum für Naturkunde in Berlin early last month, and like most paleontologists who see it, I was awestruck by its 150-million-year-old beauty. Understandably, then, the evolutionary history of birds was on my mind when – three weeks later – I watched those grackles making traces on a Georgia beach. Will Archaeopteryx trace fossils ever be found? Let’s hope so, and if they do, they deserve to be as famous as this specimen. (Photograph by Anthony Martin.)
One of the most momentous events of the post-Jurassic world happened last week with the online release of the official trailer for the upcoming movie Jurassic World. Yet within mere hours of its release, a great wailing and gnashing of teeth arose from dinosaur nerdom, as professional paleontologists and fervent paleo-fangirls and paleo-fanboys alike jumped onto it like a ravenous pack of naked, oversized, bunny-handed velociraptors (or deinonychosaurs: whatever).
Owen (Chris Pratt): “Looks like a large theropod dinosaur tooth.” Claire (Bryce Dallas Howard): “That’s not going to tell us anything. Why don’t you look at its toothmarks right next to you?” That’s just one small sample of how I would rewrite the Jurassic World script from an ichnological perspective, neatly repairing its reputation as a scientifically accurate film while retaining blockbuster entertainment value. (This still image and all others in this post were stolen shamelessly from the Jurassic World trailer.)
You see, this trailer – which lasted for all of 161 seconds – contained 257 scientific inaccuracies, which comes out to about 1.6 errors/second. OK, maybe I just made up that number, much like how some people make up movie plots, scripts, and characters. Nonetheless, the trailer had scenes depicting featherless theropods, elephant-skinned sauropods, and a non-dinosaur mosasaur that was far too big, had a frill on its back, and a non-forked tongue. It’s almost as if these were genetically recreated monsters, and not the original animals from the Mesozoic Era. Oh, the waste! Oh, the humanity!
Anyway, let’s talk about something that really matters, like traces. As far as I know – and like many others on the Internet, I plan to stay ignorant of anything that might add to my present knowledge – not one of those paleo-critics, or even the critics of the paleo-critics, mentioned the totally awesome and epic traces shown in this trailer. This egregious oversight once again reinforces my oft-asserted point that ichnology is the Rodney Dangerfield of paleontology.
So that’s why I’m here, to enlighten the masses and convert y’all to the Church of Ichnology, where there’s no tithing, dancing and drinking are required, and you can leave the church any time you feel like it. I’ve also covered this beat before, having reviewed the ichnology of Jurassic Park, which was an entire movie, not just a trailer. Even better, I know a little bit about dinosaur ichnology, having just written a book on that topic (Dinosaurs Without Bones, if you must know).
What traces in are in the trailer, you ask? The first ones shown are at 1:39, revealed with a shot panning up a concrete wall. There on the wall are sets of three more-or-less parallel scratches, some straight and some curved. The scratches vary in lengths, and a few cross-cut each another. In one set the scratches are not parallel, but form more of a fan pattern.
I knew it was worth sitting through the first 1:30 of this trailer – check out those scratch patterns!
Of course, the preceding picture means little to an ichnologist unless it has a scale. I mean, were these from Compsognathus, or something a lot bigger? Fortunately, at 1:47, Chris Pratt provides a means of comparison by placing his hand on one of the scratches, and it looks like his three middle fingers approximate the width of that mark.
So let’s assume his hand proportions are about 1.4 times the size of mine, a supposition that can only be tested by the two of us having a beer together. (Hey, it could happen.) Accordingly, we will name this conversion factor the Chris Pratt Manual Ratio™ (CPMR). My three middle fingers bunched together are 5 cm (2 in) wide, which, after multiplying it by the CPMR, would make this scraping about 7 cm (2.75 in) wide. The spaces between the scratches seem to be about twice as wide, or 15 cm (5.5 in). This would make the entire set – three scratches and the two spaces between them – 51 cm (20 in) wide. These are twice as wide as some of the biggest known theropod dinosaur tracks. Or, as I like to say whenever I encounter grizzly-bear scratches on trees: “That ain’t no squirrel.”
Put your hands on the traces, and feel their healing power and redemption! Repeat the Holy Trinity of the Church of Ichnology with me: Substrate, Anatomy, and Behavior! Amen, brothers and sisters!
Based on my detailed study of these traces for at least two minutes (perhaps less), as well as Chris Pratt’s concerned gaze following these scracthes up the wall, I am interpreting them as traces made by three claws on the feet and hands of its tracemaker, with the wider sets coming from feet and the narrower ones from hands. Overall, these traces would be anatomically appropriate for theropod dinosaurs, many of which had three digits on its feet and hands with sharp claws. Moreover, this would have been a theropod dinosaur with impressively endowed forelimbs, sufficient for helping to pull it up a wall (sorry, T. rex).
Why, that looks like an enormous three-toed foot with robust claws on their ends, and in motion as it chases the presumed protagonist of Jurassic World. Who could’ve predicted that, based on mere ichnologically based foreshadowing?
The behavior of the tracemaker can also be interpreted by looking for where the “feet” traces cross-cut the “hand” traces on the wall. This pattern would have been made by an upward movement of the tracemaker as it climbed the vertical surface. In short, these are escape traces, and they were made by a very large theropod-like dinosaur. To his credit, Chris Pratt’s character (“Owen”) totally got this.
Chris Pratt isn’t just a Guardian of the Galaxy and a dinosaur handler: he’s also an ichnologist. (By the way: what’s with the bones behind him that don’t have any toothmarks on them? And why aren’t my paleontologist friends outraged about that unforgivable error?)
Any other traces in the trailer? Oh yeah, and it’s a good one. At 1:51, Bryce Dallas Howard (“Claire”) picks up a hardhat that clearly was not hard enough to prevent serious brain leakage in its former wearer. The trace is a sharply defined gouge that nearly cleaves the hardhat into two half-hats. This trace is either from a claw or tooth, but because it’s by itself, I’m going to surmise it was from a single strategically employed claw. How wide was the claw? We can figure that out by using the Bryce Dallas Howard Pollex Ratio™ (BDHPR) of 1.0, which assumes her thumb is the same width as mine, 2.2 cm (0.9 in). (Yes, I have petite thumbs. You have a problem with that?)
Based on this unit of measurement, the split seems to be at least three times her thumb width, or minimally 6.6 cm (2.6 in) wide. Which, incidentally, is about the same width as the scratches left on the concrete wall, which I also interpreted as coming from claws, and which neatly connects the escaped “dinosaur” to this heinous act committed on what was no doubt an out-sourced employee who did not have health insurance. Coincidence? No, it’s ichnology!
Hey, this hardhat is defective! Let’s check the warranty. Yup, sure enough: “Does not cover hybrid dinosaur attacks.” But at least we got a cool trace out of it.
So despite all of the problems my paleontological colleagues justifiably noted for the dinosauroid animals depicted in the trailer, I am encouraged that Jurassic World will have enough ichnology in it to persuade me to leave a buttock-shaped impression on my theater seat in June 2015. But there had better be tracks, nests and feces in it, otherwise you’ll see my footprints going straight out of the theater.
After an undramatic (but still tiring) trip from Atlanta, Georgia to Berlin, Germany, I’m happy to be attending the Society of Vertebrate Paleontology annual meeting (SVP) in Berlin. The meeting – with talks, posters, and various social events – officially begins today (Wednesday, November 5) and continues through Saturday, November 9, but like all good paleontology meetings, it also has field trips before and afterwards.
Aside from my being with more than a thousand other paleontologists, exchanging information about the latest research, and enjoying good German beer while learning about this research (all of these are connected, I assure you), one of the main reasons why I am so far from Georgia is to present some of my research, too. It’s very much in the preliminary stages, but my coauthors and I thought it would be good to put this work out for other paleontologists to examine, poke, prod, and otherwise leave their impressions on it before we present it in a formal, peer-reviewed paper. I’ll be providing pictures and words expressing our work in a poster session today.
Series of small grooves in an Early Cretaceous (about 100 million-year-old) sandstone in Arches National Park, Utah (USA). Notice how they make radiating patterns, too. Do you wonder what made these trace fossils? If so, join the club. My coauthors and I take a semi-educated guess, which is just below for your reading pleasure. (Photograph by Anthony Martin, taken in June 2012; scale in centimeters.)
The following abstract summarizes the work, but the preceding picture might help, as does this one-sentence summary at the start of the poster:
Linear grooves in Early Cretaceous sandstone beds of the Early Cretaceous (Aptian-Albian) Cedar Mountain Formation are likely feeding trace fossils made by a beaked vertebrate, such as a pterosaur or bird.
VERTEBRATE FEEDING TRACE FOSSILS IN THE CEDAR MOUNTAIN FORMATION (LOWER CRETACEOUS), ARCHES NATIONAL PARK, UTA (USA): BIRD, PTEROSAUR, OR UNKNOWN TRACEMAKER?
MARTIN, Anthony J., Emory University, Atlanta, GA, USA, 30322; KIRKLAND, James I., Utah Geological Survey, Salt Lake City, UT, USA; MILNER, Andrew R.C., St George Dinosaur Discovery Site at Johnson Farm, St. George, UT, USA; SANTUCCI, Vincent L., National Park Service, Washington, DC, USA.
ABSTRACT
Abundant linear grooves on sandstone bedding planes of the Ruby Ranch Member of the Cedar Mountain Formation (Lower Cretaceous) in Arches National Park (Utah, USA) are interpreted as feeding traces made by a beaked vertebrate, such as a bird or pterosaur. These grooves have regular lengths (15.7 ± 2.0 mm), widths (3.4 ± 0.3 mm), and depths (1.5 ± 0.7 mm; n = 30), indicating a common origin related to the behavior and anatomy of their tracemakers. The trace fossils are either: solitary, bundled together as parallel groups of 4-8 grooves, or form semi-circular to circular patterns of 35-70. Bundles forming arc-like patterns are 13-15 cm wide. Grooves are on the same surface with runzelmarken, invertebrate trails, tridactyl theropod tracks, and a didactyl dromaeosaurid track. Forms and patterns of these features do not correspond to any known inorganic structures or invertebrate traces, nor traces made by fish. Thus they are considered as trace fossils made by either birds or pterosaurs. Runzelmarken and laminations imply that algal films bound sedimentary surfaces and helped to preserve these trace fossils and their associated theropod tracks. Hence the grooves may have been grazing traces, in which tracemakers gouged just underneath and parallel to algal films by using hard body parts, such as beaks. If so, beaks would have been 3-4 mm wide and groove lengths would have been linked to beak length and neck movement. The diameter of the semicircular and circular patterns suggests that the tracemakers were relatively small vertebrates. Arc-like patterns of clustered grooves could have been made by the tracemaker standing in one spot or shifting laterally to systematically mine the surface. However, no pes tracks were observed in direct association with these grooves. Hence the traces also may have been formed.
Many thanks to my coauthors – Jim Kirkland, Andrew Milner, and Vincent Santucci – for their help on this research, which hopefully will get a little bit of the attention it deserves here in Berlin. Stay tuned this week for more ichnologically related posts, which I’ll try to write and publish in between all of the aforementioned enjoyable exchanges and German beer.
The seagull looked peaceful on that beach, lying on its left side with its eyes closed. Yet it was a permanent quietude, as only its head was there.
This disembodied head stuck out as a white spot with a red edge, perched on top of a pile of dull-brown, dead cordgrass. The torso so recently connected to this head was nowhere to be seen, and I could find no tracks belonging to the gull or any other animal nearby. It looked as if it had been placed there as an object of art, ready for erudite admirers – wine glasses in hand – to comment on its broader themes and nuanced metaphors. To a ichnologist, though, it also spoke of a sudden death, and one likely dealt by a aerial predator.
The place where I saw this gruesome sign was on Wassaw Island, Georgia. Wassaw is the only island on the Georgia coast that was never logged or otherwise developed by European or Americans, hence it retains a more primitive feel compared to most other Georgia islands. You can only get there by boat, and in this instance our boat captain and guide – John Crawford – had taken our field-trip group there to learn about its unique natural history. Because of its intact environments and general lack of human influence on the landscape, I was not surprised to see something new on Wassaw. However, I haven’t seen anything like this since.
Within minutes of arriving on the island, this beheaded seagull presented a little mystery for us. As mentioned before, tracks and the rest of the body were not visible, nor were any droplets of blood around its head, either. Moreover, its dry feathers and the freshness of its fatal wound – a clean severing of its neck vertebrate – also meant it had not washed up on shore. Where did it die, and how did it get there?
After ruling out the land and sea, we looked above the beach, and realized that the attack must have been delivered up there, in the air. We then imagined what could have possessed the bulk, ferocity, and other means to chop through a seagull’s neck while in flight. The list of suspects was a short one, and we quickly narrowed it down to one: a bald eagle.
Our hypothesis was not so far-fetched, as bald eagles don’t just eat fish, but also kill and eat other birds, including gulls. This meant the seagull head we saw that morning was very likely a result of bird-on-bird predation. Extending this a bit further into the evolutionary pasts of these birds, it reflected a time when when their non-avian dinosaur ancestors killed and were killed by similar behaviors, but on the ground.
How did birds evolve flight from non-flighted theropod ancestors? No doubt one of many selection pressures exerted on non-avian dinosaurs was predation. Any means for increasing the likelihood of escape from predators also bestowed a greater probability for passing on genes coding for that “escaping trait” to the next generation of not-quite-flighted dinosaurs.
Of course, flight has evolved for many uses in birds. Nevertheless, making a quick getaway from mortal peril is still one of them. Yet flight has also been used as a means for enhancing predation in the birds that kill other birds, exerting new and different selection pressures on prey. This example of an evolutionary back-and-forth “arms race” between predators and prey is often nicknamed the Red Queen hypothesis, named after Lewis Carroll’s character in Alice in Wonderland. Only now I will change her line (said to a fleeing Alice) about running in place:
Now, here, you see, it takes all the running you can do to keep in the same place.
to a more avian-appropriate one:
Now, here, you see, it takes all the flying you can do to keep in the same place.
Still, In this Georgia-coast example, a more appropriate literary allusion would have been to the Queen of Hearts from Alice in Wonderland, a decapitating character famous for uttering the line, “Off with their heads!” In this sense, the Red Queen and Queen of Hearts meet in the arms race between predators and prey.
Will this “Red Queen of Hearts” scenario happen again during eagle and seagull conflicts? Yes: that is, unless the seagulls’ descendants adapt, which may be followed by the eagles’ descendants adapting to these changes. And on it goes, this evolution of the now blending with the then, a reminder that these days of the dead affect those of the living, as well as those not yet alive.
















































{kind=link}
{kind=link}
{kind=link}
{kind=link}